Evidence from homology
One of Darwin’s central arguments for the common ancestry of diverse animal species concerned the fact that similar body structures, especially skeletal features, serve different purposes in organisms that live in different environments or environmental niches. For example, many animal species have pentadactyl (five-fingered) limbs with similar bones arranged in similar ways, yet pentadactyl limbs serve widely varied purposes:
What can be more curious than that the hand of a man, formed for grasping, that of a mole for digging, the leg of the horse, the paddle of the porpoise, and the wing of the bat, should all be constructed on the same pattern, and should include the same bones, in the same relative positions?Charles Darwin, On the Origin of Species By Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life (London: Murray, 1859), 434. Available online here.
Darwin was not the first scientist to notice these similarities, of course. The ancient Greek philosopher Aristotle made many such observations, and biologists prior to Darwin were keenly aware of them. The eminent anatomist Sir Richard Owen, a contemporary of Darwin’s, coined the term homology to distinguish this type of anatomical similarity from analogy, which refers to similarities between the functions (rather than similarities between the parts) of organs or anatomical structures. For example, the flippers of a whale are analogous to the fins of a fish because they serve a similar purpose; but they are not homologous, because they are not comprised of similar parts arranged in similar ways. In contrast, the wings of a bat and the paws of a dog are not analogous, because they serve wholly different purposes; but they are homologous insofar as they consist of similar bones arranged in a similar pattern.
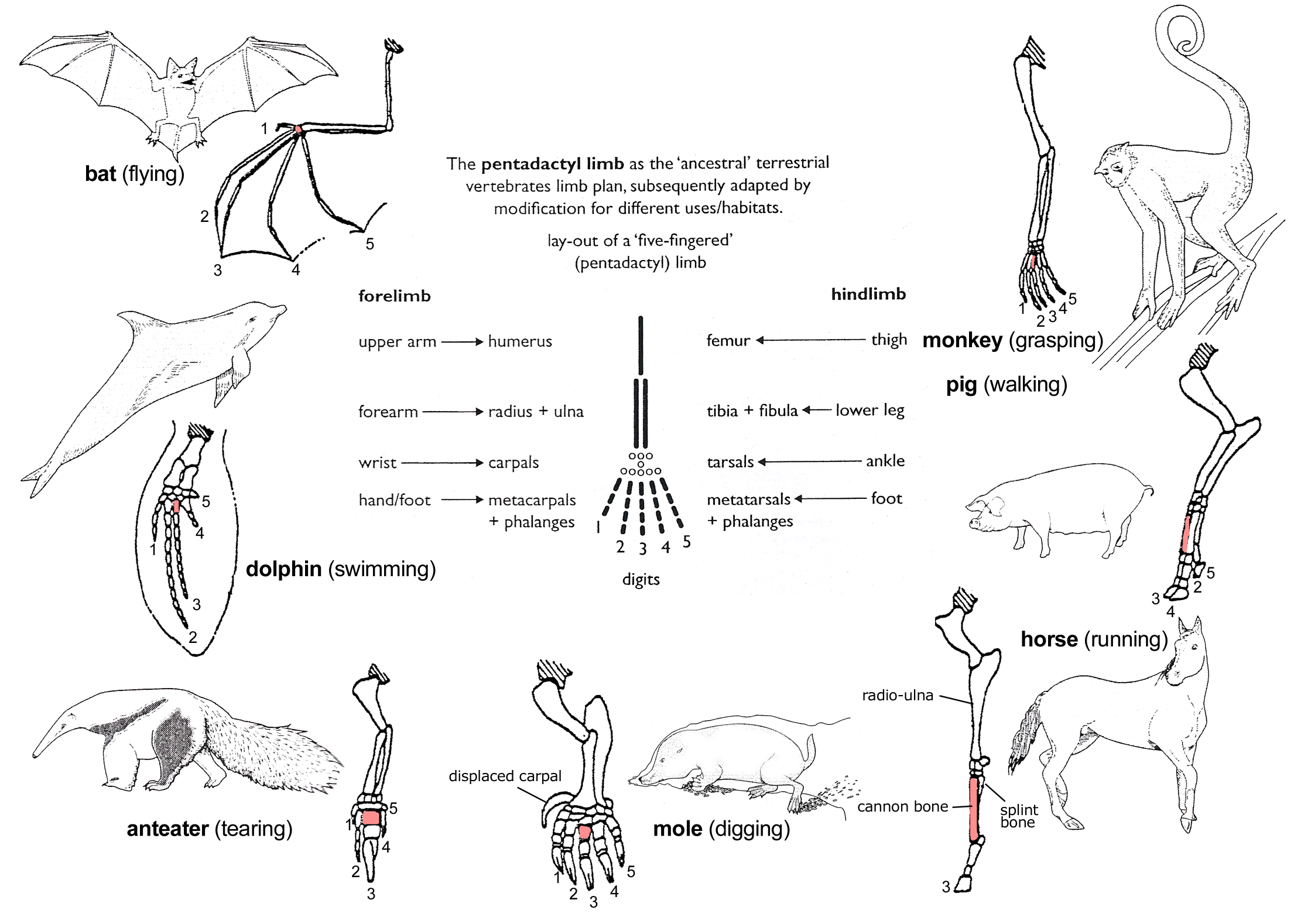
This illustration shows homologies in the limb bones of several pentadactyl species.
Like most other biologists of his time, Owen believed that God had created species separately, and he suggested that God had employed the same “archetype”—that is, a common pattern or blueprint—in designing homologous features of diverse organisms.Owen, On the Nature of Limbs (1849), available online here. Owen was not the first to suggest that God had created multiple species according to a common archetype or pattern. For further discussion of the history of Owen’s perspective, see Edward J. Larson, Evolution: The Remarkable History of a Scientific Theory (New York: Modern Library, 2004), 42-46. However, Darwin was unsatisfied with Owen’s explanation, contending that homologies could be better explained by his own theory of evolution. The reason so many animals share homologous pentadactyl limbs, Darwin argued, is that they descended from a five-fingered ancestor species, whose progeny had evolved and diversified in different environments.
Darwin’s explanation of homology is further supported by the existence of so-called vestigial structures—parts of an organism that do not appear to serve any function but are homologous to functional structures in other organisms. For example, the weight-bearing part of a horse’s hoof is homologous to the middle toe of a pentadactyl foot (with a greatly enlarged toenail); but its leg also contains shrunken and seemingly useless bone structures fused to either side. These extra toes are homologous to the second and fourth digits (index finger and ring finger) of a pentadactyl hand or foot, and are thought to be vestiges of the horse’s evolutionary ancestry. Countless additional examples of homologous structures (called homologs) and vestigial structures have been cited as evidence of common ancestry for diverse animal species.
Homology has become so foundational to modern evolutionary theory that the term homology itself has been redefined in contemporary usage to include only similarities that arise by descent from a common ancestral species.See, for example, the definition of homologous in A Dictionary of Biology, 6th edition: “Describing a character that is shared by a group of species because it is inherited from a common ancestor.” This redefinition of homology, however, invites an obvious objection. If homology refers, by definition, to similarities due to common ancestry, then we cannot determine whether something really is homologous unless we already know whether it arose by common ancestry. Given this new definition, therefore, homology cannot be cited as evidence of common ancestry, on pain of circular reasoning.For a humorously whimsical illustration of the circularity objection, see this YouTube video. The circularity objection can be avoided, of course, by sticking with the original definition of homology introduced by Richard Owen, just as Darwin did in his arguments.
On the other hand, some Christians today also stick with Owen in believing that homologies are products of God’s creative blueprints rather than products of common ancestry. The evidential significance of homology has also been disputed by some biologists who accept the idea of universal common descent. For example, biochemist Michael Denton agrees with Darwin’s claim that homology indicates common ancestry, but he rejects the Darwinian idea that homologs can be explained as the result of natural selection acting on random variations. The fact that many homologs serve diverse purposes in different organisms makes it difficult to explain how they were favored by natural selection, Denton argues. To explain the origin of the pentadactyl limb in terms of natural selection, for example, we would have to show that the five-fingered bone structure was beneficial to a specific ancestral species living in a specific environment. However, the functional versatility of the pentadactyl limb strongly suggests that it is not adapted to any specific purpose, and the same goes for many other homologs:
Many of the taxa-defining homologs and ground plans—such as the pentadactyl limb, angiosperm flower, and the floral formulae of the angiosperm sub-clades—have never been shown to have any specific utility in any actual organism in any actual environment. … If many of the homologs and ground plans are indeed non-adaptive and have never served any specific functional end, the entire Darwinian narrative is rendered a house of cards.Michael Denton, Evolution: Still a Theory in Crisis (Seattle: Discovery Institute Press, 2016), 222. Emphasis in original.
In Denton’s view, the origin and persistence of many homologs are better explained by the hypothesis that certain biological forms or structures are pre-programmed into the finely-tuned laws and properties of matter, so that they emerge almost inevitably.In his book Evolution: Still a Theory in Crisis (Seattle: Discovery Institute Press, 2016), Denton writes: “Just as a crystal of common salt arises when a solution of sodium and chloride ions evaporates, a snow crystal forms when water freezes, or a new atom arises when two nuclei collide and fuse in a stellar interior, so the basic homologs or Types (the ‘atoms’ of biology) should arise, from the cellular to the organismic level, from the self-organizing properties of particular classes of matter—more specifically, of particular classes of biomatter.” (p. 251) We’ll revisit Denton’s hypothesis in Chapter 11. A few prominent evolutionary theorists, including Richard Lewontin and Stephen Jay Gould, have similarly suggested that developmental constraints rather than functional adaptations play the central role in steering evolution toward certain forms (though Gould emphasized the role of chance and unpredictable contingencies, in contrast to Denton’s suggestion of inevitability).See Edward J. Larson, Evolution: The Remarkable History of a Scientific Theory (New York: Modern Library, 2004), 280-281 for further discussion.
So, the explanation of homology has been a topic of vigorous dispute, not only by critics of evolutionary theory but also among proponents of common descent. Nonetheless, most parties to these debates agree that homologous structures are at least partly caused—at the physiological level—by similar genetic information, especially in the gene-regulatory networks that control growth and development. The pentadactyl limb and other homologs are manifestations of underlying similarities in the genetic blueprints shared by diverse organisms. Since homologs depend on similar genes, evidence from homology should be interpreted in light of evidence from genetics. Moreover, claims about common ancestry can be corroborated or refuted by the fossil record. Thus, the evidence from homology cannot stand on its own: it must be grounded in evidence from genetics and paleontology. To those categories of evidence we now turn.